
|
Con
esta denominación común nos referimos a tres especies
de cetáceos que se distinguen osteológicamente
por presentar el cráneo extremadamente asimetrico, el
orificio respiratorio izquierdo mucho mayor que el derecho y
dientes, generalmente, solo presentes en la mandíbula.
Entre
otras muchas características la diferencia de tallas
y la proporción del tamaño de la cabeza sirven
para distinguir a dos familias. Otras
2 especies denominadas también cachalotes pertenecen
a la Familia
KOGIIDAE. El
cachalote pigmeo (Kogia
breviceps) y el
cachalote enano (kogia
sima) son mucho
más pequeños con tallas inferiores a los 4 m.
y cierto aspecto aspecto de tiburón.
La aleta dorsal esta bien definida y el tamaño de la
cabeza es proporcional. (15% de
la longitud corporal). |
|
|
|
cachalote
pigmeo ........ Kogia breviceps 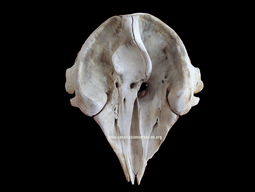  Cráneo de K breviceps (sup) y K sima (inf) |
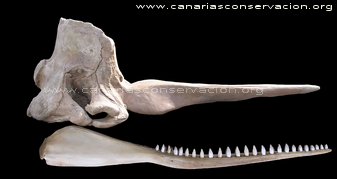 Cráneo de P macrocephalus. Macho 14,5 m Fotos y Texto ® M. Carrillo 2018 |
|
|
Catalogada como
"vulnerable" en el CEEA* y CCEP**.
|
|
